Scopriamo cos’è il “Lac Mer”, quale zona identifica e alcune informazioni utili che bisogna sicuramente conoscere
Il termine “Lac Mer” è stato utilizzato per la prima volta da Gignoux (1950) per indicare l’antico paleoambiente del Mediterraneo orientale (dal Tortoniano al Messiniano). Questo ambiente è stato creato a partire dal Tortoniano dal scollamento e di conseguenza dall’isolamento del bacino di Parathethyan rispetto al mare aperto. Pertanto, l’apporto fluviale ha gradualmente trasformato l’acqua marina in salmastra, mentre l’assemblaggio faunistico è cambiato da marina a quella marina interna e infine a quella lagunare desalinizzata (lac-mer) ed era rappresentata prima da Cerithes e poi da Congerie. Da questo tipo di fauna è derivata l’attuale fauna del Caspio.
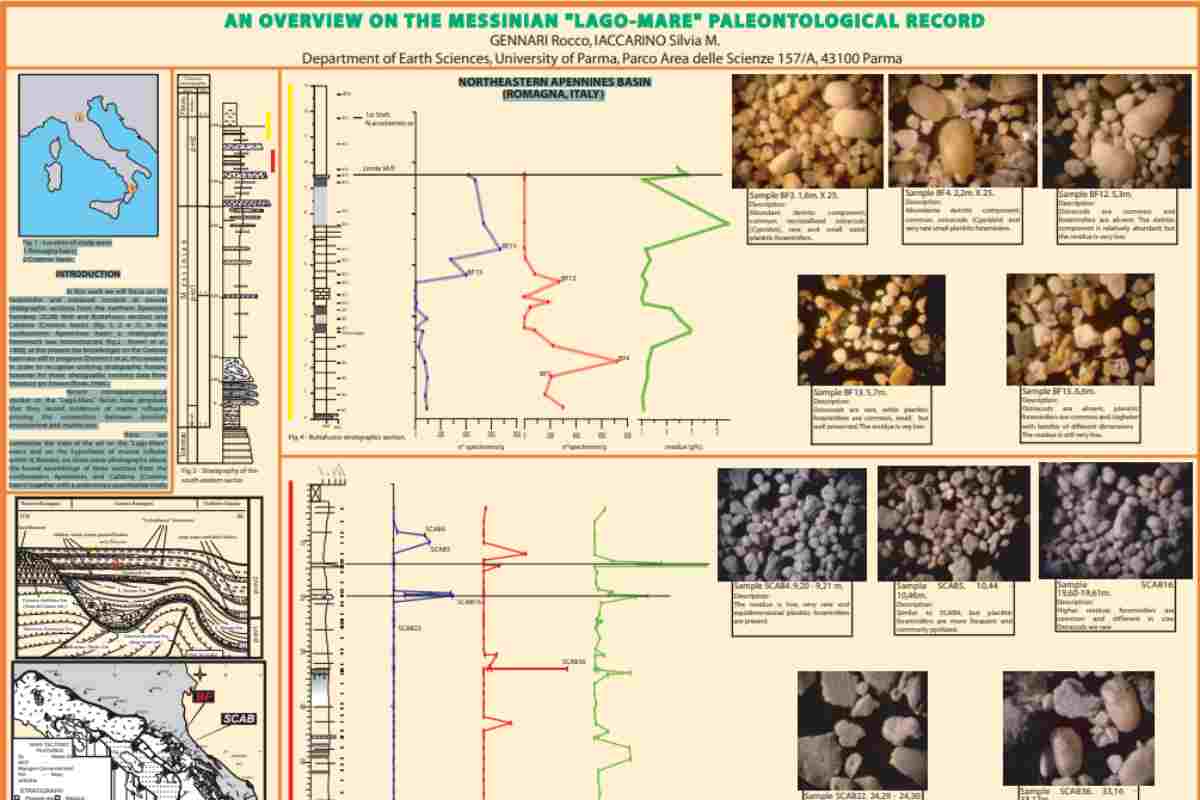
Questo scenario è stato richiamato da Ruggieri (1967) per descrivere il paleoambiente dei bacini mediterranei occidentali del Messiniano, che dopo la chiusura del collegamento atlantico si sono “trasformati in una serie di lagune, che o si sono asciugate o si sono trasformate in una serie di lagune, che si sono prosciugate o sono state gradualmente desalinizzate”. In effetti, una fauna di gasteropodi oligohaline (Melanopsis) è stata recuperata all’interno dei depositi correlativi di molti siti del Mediterraneo occidentale. Durante gli
Negli anni ’70 il Deep Sea Drilling Project fornì una nuova serie di dati e venne ipotizzata la teoria della crisi di salinità del Mediterraneo (Hsü et al., 1973). In seguito, essi proposero di utilizzare il termine “Lago-Mare” (la traduzione italiana di Lac-Mer) per traduzione italiana di Lac-Mer) per indicare “l’ultimo paleoambiente oligohalino messiniano”. Gli autori hanno collocato questo evento approssimativamente dopo la deposizione di evaporiti e prima della sedimentazione marina del Pliocene.
Sedimentazione marina del Pliocene
Secondo il modello di dessicazione (Hsu, Ryan & Cita, 1973) il Mediterraneo si è prosciugato dopo la deposizione dell’evaporite; recentemente questo evento è stato datato da Krijgsman et al. (1999) a 5,61 Ma. Sulla non conformità generata dal disseccamento si è depositata l’Unità Evaporitica Superiore e, successivamente, la Formazione di Arenazzolo. Bonaduce & Sgarella (1999) hanno distinto nei depositi “Lago-Mare” due diverse biofacies, distintamente rappresentate nelle successioni della Sicilia meridionale. La prima è caratterizzata da popolazioni monospecifiche di Cyprideis e/o da popolazioni monospecifiche di Ammonia beccarii tepida.
I molluschi ipoalini possono possono essere presenti in alcuni strati. Questa biofacies è stata interpretata come rappresentativa di un ambiente iperalino con afflusso di acqua salmastra o dolce ed è stata trovata all’interno dell’Unità Evaporitica Superiore tra il sesto e il settimo letto di gesso. La seconda biofacies, quella superiore, è caratterizzata dalla biozona Loxoconcha diaffarovi (Carbonnel, 1978) che dovrebbe corrispondere alla fauna caspio-brascosa. In Sicilia la biofacies 2 è rappresentativa della Formazione di Arenazzolo.
La seconda biofacies è ritenuta di origine paratetica (Bonaduce & Sgarella, 1999; Gliozzi, 1999). La comparsa di questo assemblaggio e il passaggio da iperalini a oligohaline o salmastre.





